Cours : Etude qualitative du modèle prédateur-proie. Oscillations du système prédateur-proie (modèle de Lotka-Voltaire) Analyse mathématique des modèles prédateur-proie
La modélisation mathématique des processus biologiques a commencé avec la création des premiers modèles simples d'un système écologique.
Disons que les lynx et les lièvres vivent dans une zone fermée. Les lynx ne mangent que des lièvres, et les lièvres - aliments végétaux, disponible en quantité illimitée. Il est nécessaire de trouver des caractéristiques macroscopiques qui décrivent les populations. Ces caractéristiques sont le nombre d'individus dans les populations.
Le modèle le plus simple de la relation entre les populations de prédateurs et de proies, basé sur l'équation de croissance logistique, est appelé (ainsi que le modèle compétition interspécifique) du nom de ses créateurs - Lotka et Volterra. Ce modèle simplifie grandement la situation étudiée, mais reste utile comme point de départ dans l’analyse du système prédateur-proie.
Supposons que (1) la population de proies existe dans un environnement idéal (indépendant de la densité) où sa croissance ne peut être limitée que par la présence d'un prédateur, (2) il existe un environnement tout aussi idéal dans lequel il existe un prédateur dont la population la croissance n'est limitée que par l'abondance des proies, (3 ) les deux populations se reproduisent continuellement selon l'équation de croissance exponentielle, (4) le taux de consommation de proies par les prédateurs est proportionnel à la fréquence des rencontres entre eux, qui, à son tour, est fonction de la densité de population. Ces hypothèses sous-tendent le modèle Lotka-Volterra.
Laissez la population de proies croître de façon exponentielle en l’absence de prédateurs :
dN/dt = r 1 N 1
où N est le nombre et r est le taux de croissance instantané spécifique de la population de proies. Si des prédateurs sont présents, ils détruisent les proies à un rythme déterminé, d'une part, par la fréquence des rencontres entre prédateurs et proies, qui augmente à mesure que leur nombre augmente, et, d'autre part, par l'efficacité avec laquelle le prédateur détecte et capture ses proies. proie lors de la rencontre. Le nombre de victimes rencontrées et mangées par un prédateur N c est proportionnel à l'efficacité de la chasse, que nous exprimerons par le coefficient C 1 ; nombre (densité) de proies N et temps passé à chercher T :
N C = C 1 NT(1)
À partir de cette expression, il est facile de déterminer le taux spécifique de consommation de proies par un prédateur (c'est-à-dire le nombre de proies mangées par un prédateur individuel par unité de temps), souvent également appelé réponse fonctionnelle du prédateur à la densité de population. de la proie :
Dans le modèle considéré C1 est une constante. Cela signifie que le nombre de proies retirées d’une population par les prédateurs augmente linéairement avec l’augmentation de la densité de population (ce qu’on appelle la réponse fonctionnelle de type 1). Il est clair que le taux total de consommation de proies par tous les individus du prédateur sera :
![]() (3)
(3)
Où R- taille de la population de prédateurs. Nous pouvons maintenant écrire l’équation de croissance de la population de proies comme suit :
En l’absence de proies, les prédateurs meurent de faim et meurent. Supposons également que dans ce cas la population de prédateurs diminuera de façon exponentielle selon l'équation :
![]() (5)
(5)
Où r2- mortalité instantanée spécifique dans la population de prédateurs.
Si des proies sont présentes, les prédateurs capables de les trouver et de les manger se reproduiront. Le taux de natalité dans une population de prédateurs dans ce modèle dépend uniquement de deux circonstances : le taux de consommation de proies par le prédateur et l'efficacité avec laquelle la nourriture consommée est transformée par le prédateur en sa progéniture. Si on exprime cette efficacité par le coefficient s, alors le taux de natalité sera :
![]()
Puisque C 1 et s sont des constantes, leur produit est également une constante, que nous notons C 2. Ensuite, le taux de croissance de la population de prédateurs sera déterminé par l'équilibre de la fécondité et de la mortalité selon l'équation :
![]() (6)
(6)
Les équations 4 et 6 forment ensemble le modèle Lotka-Volterra.
On peut étudier les propriétés de ce modèle exactement de la même manière que dans le cas de la concurrence, c'est-à-dire : en construisant un diagramme de phase sur lequel le nombre de proies est tracé le long de l'axe des ordonnées et le prédateur est tracé le long de l'axe des abscisses, et en y traçant des lignes d'isocline qui correspondent à une taille de population constante. À l'aide de telles isoclines, le comportement des populations de prédateurs et de proies en interaction est déterminé.
Pour la population de proies : d’où
Ainsi, puisque r et C 1 sont des constantes, l'isocline de la proie sera la ligne sur laquelle le numéro du prédateur (P) est constant, c'est-à-dire parallèle à l'axe des x et coupant l'axe des ordonnées au point P = r 1 / C1. Au-dessus de cette ligne, le nombre de proies diminuera et en dessous il augmentera.
Pour une population de prédateurs :
d'où
Parce que r2 et C 2 - constantes, l'isocline du prédateur sera la ligne sur laquelle le nombre de proies (N) est constant, c'est-à-dire perpendiculaire à l'axe des ordonnées et coupant l'axe des abscisses au point N = r 2 /C 2 . À gauche, le nombre de prédateurs diminuera et à droite, il augmentera.
Si l'on considère ces deux isoclines ensemble, on remarque facilement que l'interaction entre les populations de prédateurs et de proies est cyclique, puisque leurs effectifs subissent des fluctuations conjuguées illimitées. Lorsque le nombre de proies est élevé, le nombre de prédateurs augmente, ce qui entraîne une augmentation de la pression de prédation sur la population de proies et ainsi une diminution de son nombre. Cette diminution entraîne à son tour un manque de nourriture pour les prédateurs et une diminution de leur nombre, ce qui entraîne un affaiblissement de la pression de prédation et une augmentation du nombre de proies, ce qui entraîne à nouveau une augmentation de la population de proies. etc.
Ce modèle est caractérisé par ce qu'on appelle la « stabilité neutre », ce qui signifie que les populations effectuent le même cycle d'oscillations indéfiniment jusqu'à ce qu'une influence externe modifie leur nombre, après quoi les populations effectuent un nouveau cycle d'oscillations avec des paramètres différents. Pour que les cycles deviennent stables, les populations doivent influence externe s'efforcer de revenir au cycle d'origine. De tels cycles, contrairement aux oscillations neutrement stables du modèle Lotka-Volterra, sont généralement appelés cycles limites stables.
Le modèle Lotka-Volterra est cependant utile dans la mesure où il nous permet de démontrer que la principale tendance des relations prédateurs-proies se manifeste dans des fluctuations cycliques conjuguées de la taille de leurs populations.
Modèle de situation prédateur-proie
Considérons un modèle mathématique de la dynamique de la coexistence de deux espèces biologiques (populations) interagissant entre elles selon le type « prédateur-proie » (loups et lapins, brochets et carassins, etc.), appelé Volter-Lotka modèle. Il a été obtenu pour la première fois par A. Lotka (1925), et un peu plus tard et indépendamment de Lotka, des modèles similaires et plus complexes ont été développés par le mathématicien italien V. Volterra (1926), dont les travaux ont en fait jeté les bases de ce qu'on appelle écologie mathématique.
Qu'il y en ait deux espèce biologique qui vivent ensemble dans environnement isolé. Cela suppose :
- 1. La victime peut trouver suffisamment de nourriture pour survivre ;
- 2. Chaque fois qu'une proie rencontre un prédateur, ce dernier tue la victime.
Pour plus de précision, nous les appellerons carassins et brochets. Laisser
l'état du système est déterminé par les quantités x(t) Et yt)- le nombre de carassins et de brochets à la fois G. Pour obtenir des équations mathématiques décrivant approximativement la dynamique (évolution dans le temps) d’une population, nous procédons comme suit.
Comme dans le modèle de croissance démographique précédent (voir section 1.1), pour les victimes nous avons l’équation
Où UN> 0 (le taux de natalité dépasse le taux de mortalité)
Coefficient UN l'augmentation des proies dépend du nombre de prédateurs (diminue avec leur augmentation). Dans le cas le plus simple a- a - fjy (a>0, p>0). Ensuite, pour la taille de la population de proies, nous avons l'équation différentielle
Pour une population de prédateurs nous avons l'équation

Où b>0 (la mortalité dépasse le taux de natalité).
Coefficient b Le déclin des prédateurs est réduit s’il y a des proies dont se nourrir. Dans le cas le plus simple, nous pouvons prendre b - y -Sx (y > 0, S> 0). Ensuite, pour la taille de la population de prédateurs, nous obtenons l'équation différentielle

Ainsi, les équations (1.5) et (1.6) représentent un modèle mathématique du problème de l'interaction de population considéré. Dans ce modèle, les variables x,y est l'état du système et les coefficients caractérisent sa structure. Le système non linéaire (1.5), (1.6) est le modèle Voltaire-Lotka.
Les équations (1.5) et (1.6) doivent être complétées par des conditions initiales - données données des populations initiales.
Analysons maintenant le modèle mathématique construit.
Construisons le portrait de phase du système (1.5), (1.6) (au sens du problème X> 0, v >0). En divisant l'équation (1.5) par l'équation (1.6), on obtient une équation à variables séparables
En utilisant cette équation, nous avons
La relation (1.7) donne l'équation des trajectoires de phase sous forme implicite. Le système (1.5), (1.6) a un état stationnaire déterminé à partir de

A partir des équations (1.8) nous obtenons (puisque l* F 0, oui* F 0)
Les égalités (1.9) déterminent la position d'équilibre (point À PROPOS DE)(Fig. 1.6).

La direction du mouvement le long de la trajectoire de phase peut être déterminée à partir de ces considérations. Qu'il y ait peu de carassins. par ex. x ~ 0, puis d'après l'équation (1.6) y
Toutes les trajectoires de phase (sauf le point 0) courbes fermées couvrant la position d’équilibre. L'état d'équilibre correspond à un nombre constant de x" et y" de carassins et de brochets. Les carassins se multiplient, les brochets les mangent, meurent, mais leur nombre et celui des autres ne change pas. « Les trajectoires en phase fermée correspondent à des changements périodiques du nombre de carassins et de brochets. De plus, la trajectoire le long de laquelle se déplace le point de phase dépend de. conditions initiales. Considérons comment l'état change le long de la trajectoire de la phase. Que le point soit en position UN(Fig. 1.6). Il y a ici peu de carassins, beaucoup de brochets ; les brochets n'ont rien à manger, ils disparaissent progressivement et presque
disparaître complètement. Mais le nombre de carassins diminue également presque jusqu'à zéro et
seulement plus tard, quand il y avait moins de brochets que à, le nombre de carassins commence à augmenter ; leur taux de croissance augmente et leur nombre augmente - cela se produit jusqu'au point DANS. Mais une augmentation du nombre de carassins entraîne un ralentissement du processus d'extinction du shuk et leur nombre commence à augmenter (il y a plus de nourriture) - intrigue Soleil. Ensuite il y a beaucoup de brochets, ils mangent des carassins et les mangent presque tous (section CD). Après cela, les brochets recommencent à mourir et le processus se répète sur une période d'environ 5 à 7 ans. Sur la fig. 1.7 courbes qualitativement construites d'évolution du nombre de carassins et de brochets en fonction du temps. Les maximums des courbes alternent et le nombre maximum de brochets est en retard par rapport aux maximums de carassins.

Ce comportement est typique de divers systèmes type prédateur-proie. Interprétons maintenant les résultats obtenus.
Malgré le fait que le modèle considéré soit le plus simple et qu'en réalité tout soit beaucoup plus compliqué, il a permis d'expliquer certaines des choses mystérieuses qui existent dans la nature. Les histoires des pêcheurs sur les périodes où « les brochets eux-mêmes sautent entre leurs mains » sont compréhensibles ; la fréquence des maladies chroniques, etc., a été expliquée.
Notons une autre conclusion intéressante que l’on peut tirer de la Fig. 1.6. Si au point R. il y a une capture rapide des brochets (dans une autre terminologie - tir aux loups), puis le système « saute » au point Q, et un mouvement supplémentaire se produit le long d'une trajectoire fermée de plus petite taille, ce qui est intuitivement attendu. Si l'on réduit le nombre de brochets en un point R, alors le système ira au point S, et d'autres mouvements se produiront le long de la trajectoire taille plus grande. Les amplitudes des oscillations vont augmenter. C’est contre-intuitif, mais cela explique précisément ce phénomène : suite aux tirs sur les loups, leur nombre augmente avec le temps. Ainsi, le choix du moment de la prise de vue est important dans ce cas.
Supposons que deux populations d'insectes (par exemple, un puceron et une coccinelle, qui se nourrissent de pucerons) soient en équilibre naturel. x-x*,y = y*(point À PROPOS sur la fig. 1.6). Considérez l’effet d’une seule application d’un insecticide qui tue x> 0 des victimes et y > 0 des prédateurs sans les détruire complètement. Une diminution du nombre des deux populations conduit au fait que le point représentant de la position À PROPOS"sautera" plus près de l'origine des coordonnées, où x> 0, y 0 (Fig. 1.6) Il s'ensuit que sous l'action d'un insecticide destiné à détruire les victimes (pucerons), le nombre de victimes (pucerons) augmente et le nombre de prédateurs ( coccinelles) diminue. Il s'avère que le nombre de prédateurs peut devenir si petit qu'ils risquent une extinction complète pour d'autres raisons (sécheresse, maladie, etc.). Ainsi, l'utilisation d'insecticides (à moins qu'ils ne détruisent presque complètement les insectes nuisibles) conduit finalement à une augmentation de la population d'insectes dont le nombre était contrôlé par d'autres insectes prédateurs. De tels cas sont décrits dans des livres de biologie.
D'une manière générale, le taux de croissance du nombre de victimes UN dépend à la fois de "L" et de y : UN= a(x, y) (en raison de la présence de prédateurs et des restrictions alimentaires).
Avec un petit changement dans le modèle (1.5), (1.6), des petits termes sont ajoutés aux membres de droite des équations (en tenant compte, par exemple, de la compétition du carassin pour la nourriture et du brochet pour le carassin)
ici 0 f.i « 1.
Dans ce cas, la conclusion sur la périodicité du processus (le retour du système à son état d'origine), valable pour les modèles (1.5), (1.6), perd sa validité. Selon le type de petites modifications/ et g Les situations illustrées à la Fig. sont possibles. 1.8.

Dans le cas (1) l'état d'équilibre À PROPOS durable. Pour toute autre condition initiale, après un temps suffisamment long, c'est exactement ce qui s'établit.
Dans le cas (2), le système « tombe en désarroi ». L'état stationnaire est instable. Un tel système s’inscrit finalement dans une telle gamme de valeurs X et y, que le modèle cesse d'être applicable.
Dans le cas (3) dans un système avec un état stationnaire instable À PROPOS Un régime périodique s'établit au fil du temps. Contrairement au modèle original (1.5), (1.6), dans ce modèle, le régime périodique en régime permanent ne dépend pas des conditions initiales. Initialement faible écart par rapport à l’état stable À PROPOS n'entraîne pas de petites fluctuations autour À PROPOS, comme dans le modèle Volterra-Lotka, mais à des oscillations d'amplitude bien définie (et indépendante de la petitesse de l'écart).
V.I. Arnold qualifie le modèle Volterra-Lotka de rigide parce que son petit changement peut conduire à des conclusions différentes de celles données ci-dessus. Pour juger laquelle des situations illustrées à la Fig. 1.8, implémenté dans ce système, est absolument nécessaire Informations Complémentaires sur le système (sur le type de petits amendements/ et g).
Le modèle de Kolmogorov fait une hypothèse importante : puisqu'il suppose que cela signifie qu'il existe des mécanismes dans la population de proies qui régulent leur nombre même en l'absence de prédateurs.
Malheureusement, une telle formulation du modèle ne permet pas de répondre à la question autour de laquelle dernièrement Il y a beaucoup de débats, que nous avons déjà évoqués au début du chapitre : comment une population de prédateurs peut-elle exercer une influence régulatrice sur une population de proies afin que l’ensemble du système soit durable ? Nous reviendrons donc au modèle (2.1), dans lequel les mécanismes d'autorégulation (par exemple, régulation par compétition intraspécifique) sont absents dans la population de proies (ainsi que dans la population de prédateurs) ; par conséquent, le seul mécanisme permettant de réguler le nombre d’espèces incluses dans une communauté est la relation trophique entre prédateurs et proies.
Ici (ainsi, contrairement au modèle précédent, les solutions (2.1) dépendent naturellement du type spécifique de fonction trophique qui, à son tour, est déterminé par la nature de la prédation, c'est-à-dire la stratégie trophique du prédateur et la stratégie défensive du prédateur. proie. Les propriétés suivantes sont communes à toutes ces fonctions (voir Fig. I) :
Le système (2.1) a un point stationnaire non trivial dont les coordonnées sont déterminées à partir des équations
![]()
sous limitation naturelle.
Il existe encore un point stationnaire (0, 0), correspondant à l'équilibre trivial. Il est facile de montrer que ce point est une selle et que les séparatrices sont les axes de coordonnées.
L'équation caractéristique d'un point a la forme
![]()
Évidemment, pour le modèle Volterra classique.
Par conséquent, la valeur de f peut être considérée comme une mesure de l’écart du modèle considéré par rapport au modèle Volterra.
![]()
un point stationnaire est le foyer et des oscillations apparaissent dans le système ; lorsque l’inégalité opposée est satisfaite, il y a un nœud et il n’y a pas d’oscillations dans le système. La stabilité de cet état d'équilibre est déterminée par la condition
c'est-à-dire que cela dépend de manière significative du type de fonction trophique du prédateur.
La condition (5.5) peut être interprétée comme suit : pour la stabilité de l'équilibre non trivial du système prédateur-proie (et donc pour l'existence de ce système), il suffit qu'au voisinage de cet état la proportion relative de proies consommées par le prédateur augmente avec l'augmentation du nombre de proies. En effet, la proportion de proies (sur leur nombre total) consommée par un prédateur est décrite par une fonction différentiable, la condition de son augmentation (dérivée positive) ressemble à
![]()
La dernière condition prise à ce stade n'est rien de plus que la condition (5.5) de stabilité de l'équilibre. Avec continuité, cela doit également être réalisé dans un certain quartier du point. Ainsi, si le nombre de victimes dans ce quartier, alors.
Supposons maintenant que la fonction trophique V ait la forme montrée sur la Fig. 11, a (caractéristique des invertébrés). On peut montrer que pour toutes les valeurs finies (puisqu'il est convexe vers le haut)
c'est-à-dire que quelle que soit la valeur du nombre stationnaire de victimes, l'inégalité (5,5) n'est pas satisfaite.
Cela signifie que dans un système doté de ce type de fonction trophique, il n’existe pas d’équilibre stable non trivial. Plusieurs issues sont possibles : soit le nombre de proies et de prédateurs augmente indéfiniment, soit (lorsque la trajectoire passe près d'un des axes de coordonnées) pour des raisons aléatoires, le nombre de proies ou le nombre de prédateurs deviendra égal à zéro. Si la proie meurt, après un certain temps, le prédateur mourra également, mais si le prédateur meurt en premier, le nombre de proies commencera à augmenter de façon exponentielle. La troisième option - l'émergence d'un cycle limite stable - est impossible, ce qui est facilement prouvé.
En fait, l'expression

dans le quadrant positif est toujours positif, à moins qu'il n'ait la forme montrée sur la Fig. 11, a. Alors, selon le critère de Dulac, il n’y a pas de trajectoires fermées dans cette région et un cycle limite stable ne peut exister.
Ainsi, nous pouvons conclure : si la fonction trophique a la forme montrée sur la Fig. 11, et alors le prédateur ne peut pas être un régulateur qui assure la stabilité de la population de proies et donc la stabilité de l'ensemble du système dans son ensemble. Le système ne peut être stable que si la population de proies possède ses propres mécanismes de régulation internes, par exemple une compétition intraspécifique ou des épizooties. Cette option de régulation a déjà été évoquée aux §§ 3, 4.
Il a été noté précédemment que ce type de fonction trophique est caractéristique des insectes prédateurs, dont les « victimes » sont aussi généralement des insectes. D’un autre côté, les observations de la dynamique de nombreux communautés naturelles Les types « prédateurs-proies », qui incluent des espèces d'insectes, montrent qu'ils se caractérisent par des oscillations de très grande amplitude et d'un type bien spécifique.
Habituellement, après une augmentation plus ou moins progressive des nombres (qui peut se produire de manière monotone ou sous forme d'oscillations d'amplitude croissante), une forte baisse se produit (Fig. 14), puis l'image se répète. Apparemment, cette nature de la dynamique du nombre d'espèces d'insectes peut s'expliquer par l'instabilité de ce système aux nombres faibles et moyens et par l'action de puissants régulateurs intrapopulations des nombres en grand nombre.

Riz. 14. Dynamique des populations du psylle australien Cardiaspina albitextura se nourrissant d'eucalyptus. (Extrait de l'article : Clark L. R. La dynamique des populations de Cardiaspina albitextura.-Austr. J. Zool., 1964, 12, n° 3, p. 362-380.)
Si le système « prédateur-proie » comprend des espèces capables d'un comportement assez complexe (par exemple, les prédateurs sont capables d'apprendre ou les proies sont capables de trouver un abri), alors un équilibre stable et non trivial peut exister dans un tel système. Cette affirmation est prouvée tout simplement.
En fait, la fonction trophique devrait alors avoir la forme montrée sur la Fig. 11, ch. Le point sur ce graphique est le point de tangence de la droite tracée à partir de l’origine du graphique de la fonction trophique. Évidemment, à ce stade, la fonction a un maximum. Il est également facile de montrer que la condition (5.5) est satisfaite pour tous. Par conséquent, un équilibre non trivial dans lequel le nombre de victimes est plus petit sera asymptotiquement stable.
Cependant, nous ne pouvons rien dire sur la taille de la région de stabilité de cet équilibre. Par exemple, s’il existe un cycle limite instable, cette région doit alors se trouver à l’intérieur du cycle. Ou une autre option : l'équilibre non trivial (5.2) est instable, mais il existe un cycle limite stable ; dans ce cas, on peut aussi parler de la stabilité du système prédateur-proie. Depuis l’expression (5.7) lors du choix d’une fonction trophique comme la Fig. 11, in peut changer de signe en changeant à , alors le critère de Dulac ne fonctionne pas ici et la question de l'existence de cycles limites reste ouverte.

MODÈLE INFORMATIQUE « PRÉDATEUR-VICTIME »
Kazachkov Igor Alekseevich 1, Guseva Elena Nikolaevna 2
1 Université technique d'État de Magnitogorsk. G.I. Nosova, Institut de Construction, Architecture et Art, étudiante en 5ème année
2 Université technique d'État de Magnitogorsk. G.I. Nosova, Institut de l'énergie et des systèmes automatisés, candidat en sciences pédagogiques, professeur agrégé du Département d'informatique de gestion et technologies de l'information
Annotation
Cet article est consacré à un aperçu du modèle informatique « prédateur-proie ». L'étude suggère que la modélisation environnementale joue un rôle rôle énorme en recherche environnementale. Cette question est multiforme.
MODÈLE INFORMATIQUE "PRÉDATEUR-VICTIME"
Kazatchkov Igor Alekseevich 1, Guseva Elena Nikolaevna 2
1 Université technique d'État de Nosov Magnitogorsk, Institut de génie civil, d'architecture et d'art, étudiant du 5ème cours
2 Université technique d'État de Nosov Magnitogorsk, Institut d'ingénierie énergétique et de systèmes automatisés, docteur en sciences pédagogiques, professeur agrégé du département d'informatique commerciale et de technologies de l'information
Abstrait
Cet article donne un aperçu du modèle informatique « prédateur-victime ». L'étude suggère que la simulation environnementale joue un rôle important dans l'étude de l'environnement. Ce problème est multiforme.
La modélisation écologique est utilisée pour étudier notre environnement. Les modèles mathématiques sont utilisés dans les cas où il n'y a pas environnement naturel et il n'y a pas d'objets naturels, cela permet de faire une prévision de l'influence de divers facteurs sur l'objet étudié. Cette méthode assume les fonctions de vérification, de construction et d'interprétation des résultats obtenus. Sur la base de telles formes, la modélisation environnementale traite de l'évaluation des changements dans l'environnement qui nous entoure.
À l'heure actuelle, de telles formes sont utilisées pour étudier l'environnement qui nous entoure, et lorsqu'il est nécessaire d'étudier l'un de ses domaines, une modélisation mathématique est utilisée. Ce modèle permet de prédire l'influence de certains facteurs sur l'objet d'étude. À une certaine époque, le type « prédateur-proie » a été proposé par des scientifiques tels que : T. Malthus (Malthus 1798, Malthus 1905), Verhulst (Verhulst 1838), Pearl (Pearl 1927, 1930), ainsi que A. Lotka ( Lotka 1925, 1927 ) et V. Volterra (Volterra 1926). Ces modèles reproduisent le régime oscillatoire périodique qui résulte des interactions interspécifiques dans la nature.
L'une des principales méthodes de cognition est la modélisation. Outre le fait qu'il peut prédire les changements qui se produiront dans environnement, aide également à trouver la manière optimale de résoudre le problème. Les modèles mathématiques sont utilisés depuis longtemps en écologie pour établir des modèles et des tendances dans le développement des populations et aider à mettre en évidence l’essence des observations. La mise en page peut servir d'exemple comportement, objet.
Lors de la recréation d'objets en biologie mathématique, des prévisions de divers systèmes sont utilisées, des individualités particulières des biosystèmes sont prévues : la structure interne de l'individu, les conditions de survie, la constance des systèmes écologiques, grâce à laquelle l'activité vitale des systèmes est préservée .
L’avènement de la modélisation informatique a considérablement repoussé les limites des capacités de recherche. La possibilité d'une mise en œuvre multilatérale de formes difficiles qui ne permettent pas une étude analytique est apparue ; de nouvelles orientations sont apparues, ainsi que la modélisation par simulation.
Considérons ce qu'est un objet de modélisation. « L’objet est un habitat fermé où interagissent deux populations biologiques : prédateurs et proies. Le processus de croissance, d'extinction et de reproduction se produit directement à la surface de l'habitat. La proie se nourrit des ressources présentes dans l'environnement, tandis que les prédateurs se nourrissent des proies.
Dans ce cas, les ressources nutritionnelles peuvent être soit renouvelables, soit non renouvelables.
En 1931, Vito Volterra a dérivé les lois suivantes sur la relation prédateur-proie.
La loi du cycle périodique - le processus de destruction des proies par un prédateur conduit souvent à des fluctuations périodiques de la taille de la population des deux espèces, en fonction uniquement du taux de croissance des carnivores et des herbivores et du rapport initial de leur nombre. Loi de conservation des moyennes – nombre moyen de chaque type est constante, quel que soit niveau d'entrée
, à condition que les taux spécifiques d’augmentation de la population, ainsi que l’efficacité de la prédation, soient constants.
La loi de la violation des valeurs moyennes - lorsque les deux espèces diminuent proportionnellement à leur nombre, la taille moyenne de la population des proies augmente et celle des prédateurs diminue. Le modèle prédateur-proie est une relation particulière entre un prédateur et sa proie, dont les deux bénéficient. Les individus les plus sains et les plus adaptés aux conditions environnementales survivent, c'est-à-dire tout cela arrive grâce à sélection naturelle . Dans un environnement où il n'y a aucune possibilité de reproduction,
le prédateur détruira tôt ou tard la population de la proie, ce qui entraînera son extinction. Il existe de nombreux organismes vivants sur Terre qui, dans des conditions favorables, augmentent le nombre de leurs proches dans des proportions énormes. Cette capacité s'appelle : le potentiel biotique d'une espèce, c'est-à-dire une augmentation du nombre d'une espèce sur une certaine période de temps. Chaque espèce possède son propre potentiel biotique, par exemple les organismes en un an ne peuvent augmenter que de 1,1 fois, tandis que les organismes d'espèces plus petites, telles que les crustacés, etc. peuvent augmenter leur apparence jusqu'à 1030 fois, mais les bactéries sont toujours plus. Dans tous ces cas, la population augmentera de façon exponentielle.
La croissance démographique exponentielle est une progression géométrique de la croissance démographique. Cette capacité peut être observée en laboratoire chez les bactéries et les levures. Dans des conditions hors laboratoire, une croissance exponentielle peut être observée dans l’exemple des criquets ou d’autres types d’insectes. Une telle augmentation du nombre d'espèces peut être observée dans les endroits où il n'a pratiquement pas d'ennemis et où il y a plus qu'assez de nourriture. Finalement, avec l'expansion de l'espèce, après que les effectifs aient augmenté pendant une courte période, la croissance de la population a commencé à décliner.
Considérons un modèle informatique de reproduction des mammifères en utilisant comme exemple le modèle Lotka-Volterra. Laisser Dans une certaine zone, deux types d'animaux vivent : les cerfs et les loups. Modèle mathématique des changements de population dans le modèle Plateaux-Volterra :
Le nombre initial de victimes est xn, le nombre de prédateurs est yn.
Paramètres du modèle :
P1 – probabilité de rencontrer un prédateur,
P2 – coefficient de croissance des prédateurs aux dépens des proies,
d – taux de mortalité des prédateurs,
a – coefficient d'augmentation du nombre de victimes.
DANS tâche d'apprentissage les valeurs suivantes ont été fixées : le nombre de cerfs était de 500, le nombre de loups était de 10, le taux de croissance des cerfs était de 0,02, le taux de croissance des loups était de 0,1, la probabilité de rencontrer un prédateur était de 0,0026, le taux de croissance des prédateurs au détriment des proies était de 0,000056. Les données sont calculées pour 203 ans.
Nous explorons l'influence le coefficient d'augmentation des victimes pour le développement de deux populations, les paramètres restants resteront inchangés. Dans le schéma 1, on observe une augmentation du nombre de proies puis, avec un certain retard, une augmentation du nombre de prédateurs. Ensuite, les prédateurs assomment les victimes, le nombre de victimes diminue fortement et, par la suite, le nombre de prédateurs diminue (Fig. 1).

Figure 1. Taille de la population avec de faibles taux de natalité parmi les victimes
Analysons le changement de modèle en augmentant le taux de natalité de la victime a=0,06. Dans le diagramme 2, nous voyons un processus oscillatoire cyclique conduisant à une augmentation des effectifs des deux populations au fil du temps (Fig. 2).
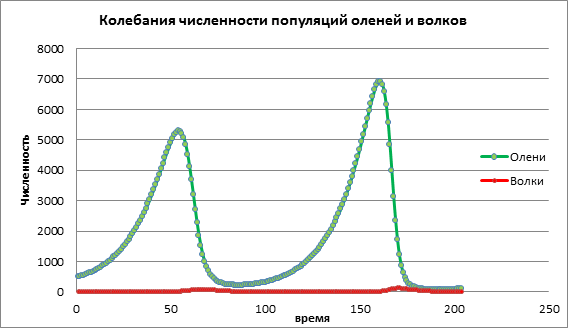
Figure 2. Taille de la population au taux de natalité moyen des victimes
Considérons comment la dynamique de la population va changer avec une valeur élevée du taux de natalité de la victime a=1,13. Sur la fig. 3, on observe une forte augmentation du nombre des deux populations, suivie par l'extinction des proies et des prédateurs. Cela est dû au fait que la population de proies a augmenté à un point tel que les ressources ont commencé à s'épuiser, entraînant l'extinction des proies. L'extinction des prédateurs est due au fait que le nombre de proies a diminué et que les prédateurs ont manqué de ressources pour survivre.
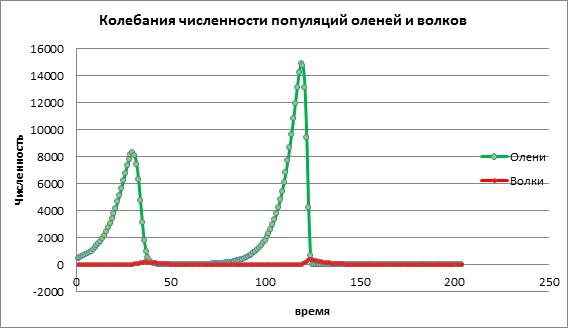
Figure 3. Taille de la population avec des taux de natalité élevés parmi les victimes
Sur la base de l'analyse des données d'expériences informatiques, nous pouvons conclure que la modélisation informatique nous permet de prédire la taille des populations et d'étudier l'influence de divers facteurs sur la dynamique des populations. Dans l'exemple ci-dessus, nous avons examiné le modèle prédateur-proie, l'influence du taux de natalité des proies sur le nombre de cerfs et de loups. Une légère augmentation de la population de proies entraîne une légère augmentation du nombre de proies qui, après un certain temps, sont détruites par les prédateurs. Une augmentation modérée de la population de proies entraîne une augmentation de la taille des deux populations. Une forte augmentation de la population de proies entraîne d'abord une augmentation rapide de la population de proies, ce qui affecte l'augmentation de la croissance des prédateurs, mais les prédateurs qui se multiplient détruisent ensuite rapidement la population de cerfs. En conséquence, les deux espèces disparaissent.
Souvent, les membres d’une espèce (population) se nourrissent de membres d’une autre espèce.
Le modèle Lotka-Volterra est un modèle d'existence mutuelle de deux populations de type « prédateur-proie ».
Le modèle prédateur-proie a été développé pour la première fois par A. Lotka en 1925, qui l'a utilisé pour décrire la dynamique des populations biologiques en interaction. En 1926, indépendamment de Lotka, des modèles similaires (et plus complexes) furent développés par le mathématicien italien V. Volterra, dont les recherches approfondies dans le domaine problèmes environnementaux a jeté les bases de la théorie mathématique des communautés biologiques ou ce qu'on appelle. écologie mathématique.
Sous forme mathématique, le système d'équations proposé a la forme :
où x est le nombre de proies, y est le nombre de prédateurs, t est le temps, α, β, γ, δ sont des coefficients qui reflètent les interactions entre populations.
Énoncé du problème
Considérons un espace clos dans lequel existent deux populations : les herbivores (« proies ») et les prédateurs. On pense qu’aucun animal n’est importé ou exporté et qu’il y a suffisamment de nourriture pour les herbivores. Alors l'équation d'évolution du nombre de victimes (uniquement victimes) prendra la forme :
où $α$ est le taux de natalité des victimes,
$x$ est la taille de la population de proies,
$\frac(dx)(dt)$ est le taux de croissance de la population de proies.
Lorsque les prédateurs ne chassent pas, ils peuvent disparaître, ce qui signifie que l’équation du nombre de prédateurs (prédateurs uniquement) devient :
Où $γ$ est le taux de perte des prédateurs,
$y$ est la taille de la population de prédateurs,
$\frac(dy)(dt)$ est le taux de croissance de la population de prédateurs.
Lorsque les prédateurs et les proies se rencontrent (la fréquence des rencontres est directement proportionnelle au produit), les prédateurs détruisent les victimes avec un coefficient ; les prédateurs bien nourris peuvent reproduire une progéniture avec un coefficient ; Ainsi, le système d’équations du modèle prendra la forme :
Solution du problème
Construisons un modèle mathématique de la coexistence de deux populations biologiques de type « prédateur-proie ».
Laissez deux populations biologiques vivre ensemble dans un environnement isolé. L'environnement est stationnaire et fournit des quantités illimitées de tout ce qui est nécessaire à la vie de l'une des espèces - les victimes. Une autre espèce - un prédateur - vit également dans des conditions stationnaires, mais se nourrit uniquement de proies. Les chats, les loups, les brochets et les renards peuvent agir comme des prédateurs, et les poulets, les lièvres, les carassins et les souris peuvent respectivement agir comme des victimes.
Pour être plus précis, considérons les chats comme des prédateurs et les poules comme des victimes.
Ainsi, les poules et les chats vivent dans un espace isolé : une cour de ferme. L'environnement offre aux poulets une nourriture illimitée et les chats ne mangent que des poulets. Notons par
$x$ – nombre de poulets,
$у$ – nombre de chats.
Au fil du temps, le nombre de poules et de chats évolue, mais nous considérerons $x$ et $y$ comme des fonctions continues du temps t. Appelons une paire de nombres $x, y)$ l'état du modèle.
Voyons comment l'état du modèle $(x, y).$ change
Considérons $\frac(dx)(dt)$ – le taux de variation du nombre de poulets.
S'il n'y a pas de chats, alors le nombre de poulets augmente, et plus vite, plus il y a de poulets. Nous supposerons que la dépendance est linéaire :
$\frac(dx)(dt) a_1 x$,
$a_1$ est un coefficient qui dépend uniquement des conditions de vie des poules, de leur mortalité naturelle et de leur taux de natalité.
$\frac(dy)(dt)$ – le taux de variation du nombre de chats (s'il n'y a pas de poulets) dépend du nombre de chats y.
S'il n'y a pas de poules, le nombre de chats diminue (ils n'ont pas de nourriture) et ils meurent. Nous supposerons que la dépendance est linéaire :
$\frac(dy)(dt) - a_2 y$.
Dans un écosystème, le taux de variation du nombre de chaque espèce sera également considéré comme proportionnel à sa quantité, mais uniquement avec un coefficient dépendant du nombre d'individus d'une autre espèce. Ainsi, pour les poulets, ce coefficient diminue avec l'augmentation du nombre de chats, et pour les chats, il augmente avec l'augmentation du nombre de poulets. Nous supposerons également que la dépendance est linéaire. On obtient alors un système d'équations différentielles :

Ce système d'équations est appelé modèle Volterra-Lotka.
a1, a2, b1, b2 – coefficients numériques, appelés paramètres du modèle.
Comme vous pouvez le constater, la nature du changement d'état du modèle (x, y) est déterminée par les valeurs des paramètres. En modifiant ces paramètres et en résolvant le système d'équations modèles, il est possible d'étudier les modèles de changements dans l'état du système écologique.
À l'aide du programme MATLAB, le système d'équations de Lotka-Volterra est résolu comme suit :
Sur la fig. 1 montre la solution du système. Selon les conditions initiales, les solutions sont différentes, ce qui correspond à différentes couleurs trajectoires.
Sur la fig. La figure 2 présente les mêmes solutions, mais en tenant compte de l'axe du temps t (c'est-à-dire qu'il y a une dépendance au temps).